- PHYLOGENÈSE
- PHYLOGENÈSELa réalité de l’évolution des êtres vivants ne se discute plus guère. Il n’en va pas de même de ses modalités. La difficulté principale réside évidemment dans l’immense durée du phénomène étudié. Peut-on espérer parvenir à déceler les mécanismes biologiques responsables des transformations qui, par exemple, à partir d’un poisson paléozoïque vieux de 350 millions d’années, ont abouti à l’Homo sapiens actuel? Cette première question conduit aussitôt à la controverse concernant les mutations, ces altérations instantanées et durables du matériel héréditaire qui se produisent aujourd’hui même avec une fréquence faible mais mesurable. Certaines mutations ont un effet considérable: ainsi, la mutation génique qui chez les Mammifères est responsable de la phocomélie (profonde modification des membres qui prennent l’aspect de nageoires), ou encore la mutation chromosomique qui chez l’homme entraîne le mongolisme. Ces deux exemples ont été repris à des auteurs qui les ont utilisés pour illustrer le rôle possible des «grosses mutations» dans l’évolution. Or, aucun Carnivore terrestre frappé de phocomélie n’a jamais pu mener la vie d’un phoque. De même ce n’est pas à partir d’un couple de Primates mongoliens que l’espèce humaine a pu se différencier. Les grosses mutations sont des monstruosités qui causent à brève échéance la mort de ceux qui les portent.On est donc réduit, paradoxalement, à ne retenir que l’accumulation des «petites mutations» pour expliquer les transformations évolutives, y compris les plus importantes. Pour ce faire, il est avantageux de se placer sur le terrain même de ceux qui refusent cette explication en distinguant dans l’évolution deux niveaux: la microévolution et la mégaévolution. La microévolution se déroule à l’intérieur de l’espèce biologique: elle entraîne la formation de races géographiques et écologiques et peut même aboutir, à la faveur d’un isolement, à la constitution d’espèces distinctes (processus connu sous le nom de spéciation). Il est largement admis que la génétique des populations rend compte convenablement de la microévolution par l’interaction des petites mutations, de la recombinaison sexuelle des génomes et de la sélection naturelle.La mégaévolution se manifeste au-delà de l’espèce biologique. Nombreux sont ceux qui doutent que la microévolution suffise à expliquer la mégaévolution et qui postulent des forces particulières et inconnues responsables de la différenciation des groupes de rang plus élevé que l’espèce.La mégaévolution se traduit par l’apparition de nouveaux groupes systématiques, par le développement de ces groupes ou phylum et éventuellement par leur extinction. Or ce vaste processus, qu’on désigne plus généralement sous le nom de phylogenèse, est strictement l’apanage des organismes à reproduction sexuée. Lorsque des êtres vivants abandonnent ce monde de reproduction, tout se passe comme s’ils s’excluaient définitivement de la mégaévolution. Cela ne suggère-t-il pas que les mécanismes élémentaires de la phylogenèse pourraient être tout simplement ceux que l’on voit à l’œuvre dans la microévolution des populations sexuées?1. L’anagenèseOn apprécie l’importance d’un groupe d’après deux critères: le degré de nouveauté de ses caractères par comparaison avec le groupe dont il est issu et le nombre des sous-groupes qu’on y reconnaît. Ainsi, parmi les Mammifères, on accorde le même rang, celui de sous-ordre, aux Pinnipèdes (de l’ordre des Carnivores) et aux Myomorphes (de l’ordre des Rongeurs): les Pinnipèdes ne comptent qu’un petit nombre de lignées, mais leur adaptation très poussée à la vie aquatique les oppose nettement à leurs ancêtres, les Fissipèdes ou Carnivores terrestres; au contraire, les Myomorphes diffèrent assez peu des Rongeurs primitifs, mais regroupent un très grand nombre de lignées. D’une façon générale, la phylogenèse combine deux processus distincts: l’apparition et la transformation des caractères ou anagenèse , la multiplication du nombre des lignées et des groupes ou cladogenèse (fig. 1).Continuité de l’évolutionL’anagenèse procède-t-elle par bonds? La paléontologie peut suggérer cette idée, car il n’est pas rare que dans une même séquence stratigraphique on voie se succéder des fossiles très différents sans aucun intermédiaire morphologique connu. Mais de tels hiatus peuvent avoir des causes variées: lacune de sédimentation, absence de fossilisation, substitution d’une espèce à une autre par compétition ou par suite d’un changement de milieu. Ce dernier événement peut aussi provoquer dans une espèce un changement morphologique non héréditaire, qui, quelle que soit sa magnitude, ne peut pas être tenu pour une véritable évolution. Ces hiatus de diverses sortes sont bien réels et compliquent suffisamment la tâche de l’évolutionniste, mais il s’y ajoute des discontinuités subjectives qui découlent de la façon dont certains paléontologistes conçoivent l’espèce. Par conviction, par nécessité ou par tradition, beaucoup d’entre eux s’en tiennent à l’espèce typologique décrite d’après quelques individus «typiques», voire un seul. Les fossiles qui s’écartent de la morphologie du «type» sont alors baptisés «formes aberrantes» ou pris pour types de nouvelles espèces. De cette façon, on perd de vue l’espèce biologique toujours représentée par des populations au sein desquelles les individus, tous aussi représentatifs les uns que les autres, se répartissent dans un champ plus ou moins étendu de variations morphologiques. L’approche typologique conduit, lorsque les fossiles récoltés sont peu nombreux, à définir «l’espèce» sur une assise trop étroite, et, lorsqu’ils sont abondants, à découper une population unique en de nombreuses «espèces» artificielles. Dans les deux cas, la comparaison des fossiles de deux niveaux successifs ne peut faire apparaître aucun lien direct entre les espèces.Inversement, chaque fois que l’approche populationnelle a été adoptée en paléontologie, il a été possible de mettre en évidence des lignées à évolution continue (fig. 2). Mais il faut bien souligner le sens particulier que prend ici le mot «continu». Les comparaisons que l’on fait entre fossiles de niveaux stratigraphiques successifs sont statistiques. On calculera par exemple la taille moyenne , en tenant compte de tous les individus adultes récoltés dans un gisement, et on exprimera la variabilité de ce caractère par un paramètre tel que l’écart type. On procédera de même dans un second gisement de niveau différent. Si les tailles moyennes de ces deux populations successives présentent une différence significative du point de vue statistique, on sera fondé à affirmer qu’il y a eu évolution. Si, en outre, cette différence, quoique significative, paraît petite par rapport à la variabilité de chacune des populations, on exprimera ce fait en disant qu’il y a eu évolution continue. On connaît maintenant de nombreux exemples de telles évolutions continues permettant de relier, par l’intermédiaire de populations successives, une espèce ancestrale à une espèce descendante très différente. Cette façon de s’exprimer n’est d’ailleurs pas entièrement satisfaisante: d’un ancêtre à son descendant, aussi différent soit-il, n’est-ce pas en quelque sorte la même espèce qu’on suit? Il doit donc être bien clair que l’objet naturel ici considéré est la lignée spécifique. Ce n’est que pour la commodité du langage qu’on donne arbitrairement un nom spécifique différent à des segments successifs d’une telle lignée.L’évolution orientéePuisque l’évolution ne procède jamais par bonds, toute transformation demande un certain temps au cours duquel les changements successifs se font tous dans le même sens. À cette évolution orientée, on donne parfois le nom d’«orthogenèse», mot superflu et même dangereux par tout ce qu’il sous-entend: irréversibilité, régularité, finalité. L’évolution est certes irréversible en ce sens qu’on ne verra jamais une transformation importante s’inverser dans tous ses détails pour aboutir à un descendant parfaitement identique à son lointain ancêtre; non pas que la chose soit interdite par quelque loi biologique, mais simplement parce que le processus anagénétique comprend de nombreux aléas dont la répétition exacte en ordre inverse ou direct est improbable. Mais ce qui est vrai de l’organisation totale d’un être vivant ne l’est pas d’un caractère partiel simple, comme par exemple la taille. Contrairement à ce qui a été longtemps admis, il n’y a pas de «loi de l’augmentation de la taille». L’exemple ultraclassique de l’évolution des Équidés, indûment invoqué à l’appui de l’orthogenèse, montre clairement que dans une même lignée la taille peut croître puis décroître, et qu’elle peut aussi ne pas varier pendant de longues durées.Les taux d’évolutionIl est difficile d’apprécier la régularité de l’évolution en se référant directement à l’écoulement du temps géologique mesuré en millions d’années, car les datations «absolues» des gisements fossilifères sont en nombre insuffisant et souvent imprécises. Mais en comparant des lignées les unes aux autres, il est facile de vérifier que l’évolution est irrégulière. Si chaque lignée évoluait à taux constant en fonction du temps absolu, on pourrait utiliser l’anagenèse d’une lignée particulière comme échelle de temps. Par rapport à cette échelle, les autres lignées évolueraient selon une loi linéaire. Ce n’est pas du tout ce qu’on observe. Le taux d’évolution d’une lignée varie donc d’une époque à l’autre.Le cheval actuel mesure au garrot environ un mètre de plus que son ancêtre l’éohippus (ou Hyracotherium ) du début de l’ère tertiaire (face=F0019 漣 50 millions d’années). Le taux moyen de cette anagenèse a donc été de 2 cm par 106 ans (fig. 3). Mais, pendant les vingt premiers millions d’années, le taux a été pratiquement nul et, au cours des derniers dix mille ans, il a même été négatif. C’est il y a vingt-cinq à trente millions d’années qu’il a connu ses valeurs les plus fortes: 20 cm, c’est-à-dire environ 20 p. 100 d’augmentation de la taille, par 106 ans. C’est là un taux élevé pour une évolution naturelle, mais extrêmement bas par comparaison avec les évolutions des races d’animaux domestiques et de plantes cultivées dues à la sélection artificielle. Admettons qu’il ait fallu 500 ans à l’homme pour obtenir à partir des chevaux normaux une race de poneys de la taille de l’éohippus. Cela donne un taux d’évolution de 2 cm par 10 ans, dix mille fois plus élevé en valeur absolue que celui de la lignée éohippus-cheval au moment de son évolution la plus rapide!On peut donc dire que l’évolution naturelle est lente: lente au regard des potentialités de l’espèce. Ce fait peut s’interpréter d’une première manière: dans la nature, les espèces sont assez bien adaptées à leur environnement et sont donc soumises à une pression de sélection beaucoup plus stabilisatrice que modificatrice. Cependant, cette interprétation ne peut s’appliquer à tous les cas, car, à chaque époque géologique, des espèces nombreuses se sont éteintes, ce qui veut dire qu’elles n’étaient pas adaptées à leur nouvel environnement. Il faut donc introduire une seconde interprétation s’appliquant aux espèces en voie d’extinction: les espèces ne supportent pas une pression de sélection modificatrice trop intense. L’observation des «races artificielles» lâchées dans la nature confirme cette façon de voir: dans la plupart des cas, elles disparaissent très rapidement; dans les autres cas elles retournent, en quelques générations, au «type sauvage». C’est que ces races sont des monstruosités entretenues par l’homme: quelques-uns de leurs «caractères partiels simples», voire un seul, ont été fortement modifiés sans égard pour le reste de l’organisme.Dans l’évolution naturelle, il n’en va pas de même: jamais un caractère ne se modifie seul. Ainsi l’augmentation de la taille chez les Équidés a été accompagnée d’une profonde réorganisation des mécanismes locomoteurs. On ne dit pas toute la vérité en présentant le développement évolutif de la patte du cheval comme une adaptation à la course. Le cheval ne court pas plus vite que le loup et l’éohippus, qui avait la conformation corporelle de ce dernier, avait comme lui la vélocité d’un coursier moderne. Mais un éohippus ou un loup qui aurait la taille d’un cheval ne tiendrait pas sur ses pattes: trois fois plus haut et vingt-sept fois plus lourd, il ferait supporter à ses os, à ses tendons et à ses souples articulations des tensions neuf fois plus fortes. Grâce à l’acquisition d’un tout autre modèle de patte, beaucoup plus rigide, le cheval est resté apte à la course malgré son augmentation de taille. On voit par l’exemple de la coaptation nécessaire des membres et de la taille (nécessité parmi d’autres) que la pénétration d’un petit herbivore dans le domaine adaptatif des Ongulés de grande taille n’est pas une mince affaire. L’évolution d’un seul caractère ne peut se faire sans un réajustement de tout l’organisme par le truchement de son génome, ce qui demande du temps.2. La cladogenèseLes taux taxonomiquesDans la mesure où la classification utilisée respecte la phylogénie réelle, tous les représentants actuels d’un groupe zoologique ou botanique descendent d’une unique espèce, qui est l’ancêtre commun du groupe. Cela ne donne qu’une idée incomplète de la cladogenèse qui s’est produite dans la descendance de cet ancêtre, car certaines lignées descendantes se sont éteintes à diverses époques géologiques. Ainsi, les Périssodactyles comptent trois familles vivantes (Équidés, Tapiridés, Rhinocérotidés) et plusieurs autres qui ont disparu au cours du Tertiaire. Les familles actuelles sont elles-mêmes appauvries: celle des Équidés se limite maintenant aux quelques espèces du genre Equus (cheval, onagre, âne, zèbre), alors qu’elle comprenait une dizaine de genres à l’époque miocène.Seule la paléontologie permet donc de connaître l’ampleur totale de la cladogenèse dans un groupe. Elle permet d’ailleurs de préciser le rythme de ce processus. On calcule pour cela des taux d’évolution d’un type particulier: alors que les taux d’évolution anagénétique étaient des taux de transformation morphologique, les taux d’évolution cladogénétique seront des taux d’apparition de sous-groupes ou taux taxonomiques (fig. 4). Un des résultats les plus constants de cette analyse est que la cladogenèse procède à un taux extrêmement variable. Une phase de cladogenèse particulièrement intense se produit en général au tout début de l’histoire du groupe, suivie d’une phase plus longue où l’anagenèse devient le processus le plus voyant.Cladogenèse et spéciationLe processus cladogénétique élémentaire est la spéciation ou subdivision d’une espèce biologique en deux espèces distinctes ou davantage. Cependant, les taux de cladogenèse que l’on calcule ne doivent pas être confondus avec des taux de spéciation: ils ne tiennent pas compte de toutes les spéciations qui se sont produites dans un groupe, mais seulement de celles qui ont donné naissance à des lignées ayant chacune une descendance, ou clade , suffisamment durable et reconnaissable pour constituer un sous-groupe distinct. On connaît des groupes, comme celui des drosophiles, à taux de spéciation élevé et à taux de cladogenèse très faible: le nombre des espèces qui apparaissent est compensé par celui des espèces qui s’éteignent, et les clades qui s’établissent sont peu différents de ceux qui les ont précédés. D’autres groupes, comme celui des Carnivores, ont un taux de spéciation beaucoup plus bas, mais ont engendré des clades originaux tout au long du Tertiaire.Il faut mentionner ici un cas particulier, la spéciation dite abrupte, qui peut se produire à la suite d’une hybridation. Les hybrides de deux espèces bien distinctes sont souvent stériles, car la trop grande dissemblance des chromosomes des deux parents perturbe leur appariement au moment de la formation des cellules sexuelles (gamètes), qui de ce fait sont anormales. Cet obstacle à la formation de gamètes féconds peut être surmonté si l’hybridation s’accompagne d’une tétraploïdie: dans ce cas, l’hybride possède une paire de chacun des chromosomes légués par ses parents, ce qui permet un appariement convenable. Mais alors, par comparaison avec les espèces parentales, l’hybride apparaît comme un tétraploïde, produisant des gamètes diploïdes. Cela isole sexuellement l’hybride et lui permet d’être le point de départ d’une nouvelle espèce et, éventuellement, d’un nouveau clade. Ce processus a joué un rôle non négligeable dans l’évolution des végétaux.3. La radiation adaptativeAnagenèse et cladogenèse, séparées jusqu’ici pour les besoins de l’exposé, sont intimement associées dans la phylogenèse selon des modalités à première vue fort variées. On peut cependant les ramener toutes à un même modèle, celui de la radiation adaptative, que l’on va définir progressivement à l’aide d’exemples.La zone adaptativeLa visite de Darwin aux îles Galapagos (dans l’océan Pacifique, à 1 000 km au large de l’équateur) fut une étape décisive dans l’histoire du transformisme. Les représentants de la famille des Fringillidés (pinsons) dans cet archipel et celui des Cocos n’ont cessé depuis lors de servir à l’illustration du fait évolutif (fig. 5). Ils descendent tous d’une même espèce venue du continent. Mais alors que les Fringillidés continentaux du monde entier ont le même type adaptatif (gros bec de granivore, pattes de passereau se posant au sol, etc.), ceux des Galapagos ont des modes de vie très divers se reflétant particulièrement bien dans la forme de leur bec: pinsons granivores terrestres normaux, pinsons mangeant la chair des cactus, pinsons arboricoles insectivores de type «mésange», pinsons de type «pic», etc. La cladogenèse du groupe des pinsons de Darwin a donc été accompagnée d’une anagenèse orientée différemment selon les lignées et qui a abouti à une divergence adaptative de ces lignées. Par ailleurs, chaque lignée a réalisé un type adaptatif connu dans d’autres groupes, ce qu’on appelle une convergence . Remarquable est la convergence du «pinson-pic» qui se réalise, non par allongement du bec comme chez le vrai pic, mais par utilisation d’une épine végétale, sorte de bec artificiel permettant de déloger les insectes cachés dans les écorces.L’espèce de pinson qui a fondé un nouveau groupe aux Galapagos a poursuivi sur le continent une évolution sans éclat. Il est facile d’imaginer pourquoi: dans la faune continentale, riche et diversifiée, les rôles adaptatifs sont distribués depuis longtemps et chaque espèce occupe une niche écologique strictement délimitée par les autres espèces. Par contre, dans l’archipel, presque inaccessible et fraîchement sorti des flots (il s’agit de volcans), il n’y avait aucun oiseau terrestre et le pinson pionnier a pu d’emblée élargir sa niche écologique. Des races géographiques et des races écologiques sont apparues, bientôt parvenues au rang d’espèces, c’est-à-dire devenues génétiquement indépendantes et capables dès lors de glisser chacune vers une niche écologique qu’elles ont interdite aux autres. C’est ainsi que les pinsons de Darwin ont conquis des niches écologiques dont l’ensemble constitue la zone adaptative de leur radiation. On voit par cet exemple que le fait, pour une espèce, d’engendrer ou de ne pas engendrer une radiation adaptative dépend de l’environnement dans lequel elle est placée.L’adaptation cléLes Rongeurs constituent parmi les Mammifères un ordre des plus prospères. L’ampleur de leur radiation est facile à mettre en évidence sur le plan des adaptations locomotrices: quelles qu’aient été les qualités du Paramys (l’ancêtre commun de tous les Rongeurs), il ne pouvait être à la fois sauteur comme la gerboise, fouisseur comme le rat-taupe, nageur comme le castor, planeur comme l’écureuil volant, etc.Le Paramys est apparu il y a cinquante millions d’années dans un monde déjà dominé par les Mammifères et où la concurrence ne faisait pas défaut. Or, presque aussitôt, il s’est répandu sur tous les continents accessibles et, en quelques millions d’années, une douzaine de sous-ordres en sont issus, dont la plupart ont survécu jusqu’à maintenant. La raison de ce succès apparaît clairement d’après la nature de ses victimes: l’expansion des Rongeurs au cours de l’Éocène coïncide avec le déclin puis la disparition de tous les Mammifères primitifs (Multituberculés, Insectivores, Prosimiens) qui utilisaient leurs incisives pour ronger. Cela indique déjà que «l’incisive faite pour ronger» est bien l’adaptation clé du groupe des Rongeurs.Tous les animaux qui utilisent ainsi leurs incisives les usent fortement et sont donc soumis à une pression de sélection en faveur de la prolongation de la croissance de ces dents au cours de la vie individuelle. Il en résulte une anagenèse qui s’observe dans les lignées des victimes de la compétition avec les Rongeurs. Mais seuls ces derniers prolongent la croissance de leurs incisives au point de la rendre permanente. En fait, toutes les lignées en compétition se rapprochaient de ce seuil, mais la première qui l’a atteint a barré l’avenir des autres: l’incisive à croissance permanente a rendu la lignée du Paramys compétitive en dehors de sa niche écologique initiale et, de proche en proche, ses descendants ont conquis une zone adaptative de plus en plus étendue. Dans cet exemple, ce qui déclenche la radiation adaptative, c’est le franchissement d’un certain seuil d’efficacité au cours du développement de l’adaptation clé.Le parallélismeL’adaptation clé s’est développée de façon lente et continue, comme dans toute anagenèse, avant d’atteindre le seuil de déclenchement de la radiation. Mais elle était encore perfectible et ne s’est pas arrêtée sur ce seuil. Ainsi s’explique de façon très simple le phénomène du parallélisme (acquisition, par différentes lignées apparentées, d’un caractère que leur ancêtre commun ne possédait pas). Ce phénomène a beaucoup dérouté les évolutionnistes; ils y ont souvent vu un argument en faveur de l’orthogenèse.Chez les Rongeurs, un bel exemple de parallélisme morphologique est celui des Hystricomorphes (mot dérivé de Hystrix , le porc-épic). Ces Rongeurs possèdent un foramen infraorbitaire extraordinairement élargi par rapport à ce qui s’observe chez d’autres Rongeurs, dont le Paramys , et chez tous les Mammifères autres que les Rongeurs. L’originalité de ce caractère a conduit naturellement les zoologues à conclure que les Hystricomorphes forment un groupe homogène. Or, par la suite, la paléontologie a établi que ce caractère s’était développé dans au moins quatre clades distincts. Le caractère hystricomorphe est un grade évolutif qui recoupe plusieurs clades. On dit encore que le groupe des Hystricomorphes correspond à une classification horizontale et que ce groupe est polyphylétique . La taxonomie moderne botanique et zoologique vise à se rapprocher le plus possible d’une classification verticale où tout groupe nommé serait monophylétique et correspondrait donc à un clade. Mais les parallélismes sont difficiles à démasquer, surtout dans les groupes qui ne se prêtent pas à l’étude paléontologique.L’étrange parallélisme des Hystricomorphes s’éclaire lorsqu’on examine la fonction du foramen infraorbitaire. Sous sa forme primitive, ce conduit ne sert qu’à quelques vaisseaux et nerfs peu importants, tandis que chez ces Rongeurs il livre passage à un gros faisceau musculaire qui rattache l’arrière de la mandibule à la région du museau. L’allongement de ce faisceau facilite beaucoup les mouvements longitudinaux de la mandibule qui, alternativement, permettent l’action des incisives ou des molaires. Il s’agit d’une coaptation entre la musculature masticatrice et les incisives développées des Rongeurs, donc d’un perfectionnement de l’adaptation clé. Chez d’autres Rongeurs, appelés Sciuromorphes (mot dérivé de Sciurus , l’écureuil), l’extension vers l’avant du faisceau musculaire ne s’est pas faite au travers du foramen infraorbitaire, mais plus extérieurement, ce qui a entraîné son oblitération complète. Cela montre que le parallélisme adaptatif ne se traduit pas obligatoirement par un parallélisme morphologique.4. L’histoire phylogéniqueL’exemple des Rongeurs ayant complété celui des pinsons de Darwin, on voit que la première condition nécessaire pour qu’une radiation adaptative se produise est qu’une espèce possède une adaptation clé utilisable dans d’autres niches écologiques que la sienne. Si cette espèce rencontre un ensemble de telles niches écologiques dépourvues d’occupant, une radiation adaptative se déclenche quel que soit le degré de perfectionnement de l’adaptation clé. Si, par contre, toutes ces niches écologiques sont occupées, leur conquête ne pourra se faire que si l’adaptation clé est suffisamment développée.Toute l’histoire évolutive des êtres vivants peut être regardée comme une succession de radiations adaptatives d’ampleurs variables. Certaines radiations se produisent à l’intérieur d’une radiation plus vaste: ainsi la radiation des Castoridés, fondée sur une adaptation à la vie aquatique, se développe à l’intérieur de la radiation des Rongeurs, puisque les Castoridés continuent à dépendre de leurs incisives à croissance permanente. Mais d’autres radiations s’accompagnent d’un changement de zone adaptative par abandon d’une ancienne adaptation clé au profit d’une nouvelle: ainsi les Insectivores, dont la radiation est fondée sur une adaptation à la capture et à l’ingestion de petites proies, comptent, dans leur descendance, la grande radiation des Ongulés, strictement adaptés au régime végétarien.Origine des adaptationsCe qui peut encore paraître obscur dans cette vision d’ensemble, c’est l’origine des nouvelles adaptations, ou plutôt l’apparition des fonctions et des organes sur lesquels reposent ces adaptations. Il semble que l’on soit enfermé dans un cercle vicieux: une nouvelle fonction nécessite un organe préexistant, mais un organe ne se développe sous la pression de la sélection naturelle que s’il a une fonction.Toute nouvelle fonction utilise effectivement un organe préexistant et, en ce sens, il est parfaitement exact de dire qu’une adaptation nouvelle suppose une préadaptation. Ainsi le vol ramé de l’oiseau, si différent de celui des autres vertébrés volants actuels (chauve-souris) ou fossiles (ptérodactyles) et tellement plus efficace, est lié à un certain type de plumes de l’aile, les rémiges. Le vol de l’oiseau n’a pas fait naître les plumes. Celles-ci préexistaient et assuraient une autre fonction, l’isolation thermique du corps, fonction qui a d’ailleurs été conservée par une partie du plumage. C’est cette fonction d’isolation qui a entraîné la différenciation des écailles de certains reptiles (écailles qui sont encore présentes, indifférenciées, sur la patte de l’oiseau) jusqu’à les transformer en plumes. Ainsi une nouvelle fonction en relaie toujours une autre sur un organe préexistant et entraîne à son tour une nouvelle différenciation de cet organe, ce qui peut fournir l’occasion d’une nouvelle adaptation, et ainsi de suite.L’homme et l’évolutionL’évolution a-t-elle un sens? Répondre oui à cette question, c’est toujours affirmer d’une manière directe ou détournée une position anthropocentrique, puisque les transformations que l’on souligne sont celles qu’on observe dans l’ascendance de l’homme: accroissement de la mobilité, de la «liberté», de la complexité, voire de la «complexité-conscience», montée du psychisme, etc. C’est faire peu de cas de puissantes radiations adaptatives, comme celle des végétaux que certains zoologistes ont qualifiée de «vaste évolution régressive» pour la seule raison que l’ancêtre commun des animaux et des plantes devait être un protiste doué de quelque motilité!Qu’il y ait eu des types d’organisation de plus en plus complexes dans de nombreuses lignées, c’est l’évidence même. Mais il faut se garder des prédictions a posteriori. La rémige de l’oiseau n’est pas en projet dans l’écaille du premier reptile. L’homme n’est en projet ni dans la première cellule vivante, ni même dans le premier primate. Chaque radiation adaptative résulte d’un concours de circonstances génétiques (ce que la lignée initiale apporte avec elle) et environnementales (ce que le milieu, y compris les autres êtres vivants, lui offre ou lui oppose) dont le caractère fortuit doit être reconnu: pouvait-on raisonnablement prévoir que l’écaille reptilienne, organe de protection mécanique, se transformerait en organe du vol ramé grâce à sa transformation préalable en organe de l’isolation thermique?Comme toujours, le point de vue anthropomorphique fausse l’étude scientifique, y compris celle de l’homme lui-même. À vouloir reconnaître une tendance privilégiée dans le mouvement général de l’évolution, à refuser de considérer l’homme comme un aboutissement, parmi d’autres, de cette évolution, on se prive de bien voir l’essentiel: à quel point l’apparition de l’homme marque une rupture dans un processus jusque-là continu. L’évolution de tous les groupes, quelle que soit la simplicité ou la complexité de leur organisation, s’est déroulée selon les mêmes modalités et en mettant en jeu les mêmes mécanismes biologiques. Par contre, le développement de l’adaptation clé de l’homme, son immense aptitude à apprendre fondée sur une fonction, l’intelligence, et un organe, le cerveau, a des conséquences originales: d’une part il y a radiation adaptative maximale dans la mesure où l’homme balaie progressivement (mais à quelle énorme vitesse à l’échelle des temps géologiques) la niche écologique de tous les autres êtres vivants, qui se trouveront bientôt tous ou éliminés ou réduits à l’état de commensaux; d’autre part il n’y a pas la moindre radiation si l’on considère que l’espèce humaine n’est pas en voie de se subdiviser, mais bien au contraire de fondre les races qui témoignent encore des stades primitifs de son expansion géographique.Au cours du Quaternaire, l’évolution biologique de l’homme, manifestée essentiellement par l’accroissement du volume de son cerveau, a progressé à un taux parmi les plus élevés que l’on connaisse en paléontologie, ce qui indique l’importance prise depuis plus d’un million d’années par l’intelligence dans l’adaptation de l’homme à son environnement. En même temps que cette fonction se développe, elle rend possible un autre type d’évolution, l’évolution culturelle , qui tout d’abord est lente et paraît seulement refléter l’évolution biologique des Australopithèques et de leurs descendants les Pithécanthropes. Mais ensuite l’évolution culturelle s’accélère d’elle-même, comme en témoigne la succession de plus en plus rapide des «industries» préhistoriques, puis l’histoire des civilisations.Puisque l’évolution biologique passée s’est faite sans projet, on ne peut lui donner un sens qu’en se plaçant sur un plan métaphysique. Il n’y a cependant rien de métaphysique dans l’idée que l’évolution future pourrait avoir un sens, celui que l’homme, seul être capable de faire des projets, lui donnerait. Cela suppose toutefois que l’homme parvienne d’abord à maîtriser les processus de l’évolution non biologique (technique, économique, sociale, politique, etc.), dont il paraît à l’heure actuelle être le jouet et tout le monde vivant avec lui.
phylogenèse [ filoʒənɛz ] n. f.• 1874; d'apr. l'all., du gr. phulon « race » et -genèse♦ Biol. Histoire évolutive des espèces, des lignées et des groupes d'organismes (par oppos. à ontogenèse).
● phylogénie ou phylogenèse nom féminin (du grec phûlon, tribu) Succession des espèces animales ou végétales que l'on suppose descendre les unes des autres et qui constituent un phylum. ● phylogénie ou phylogenèse (difficultés) nom féminin (du grec phûlon, tribu) Orthographe Attention au y du préfixe phylo- (du grec phÛlon, tribu, espèce ; sans rapport avec philo-, du grec philos, ami). Attention également au e de -genèse, qui devient é dans les dérivés phylogénétique, phylogénie, phylogénique.phylogenèse ou phylogénèse ou phylogénien. f. BIOL Modèle de la formation des espèces par évolution d'espèces ancestrales.|| Science qui étudie cette évolution et définit les phylums.⇒PHYLOGENÈSE, PHYLOGÉNIE, (PHYLOGENÈSE, PHYLOGÉNÈSE)subst. fém.BIOL. [P. oppos. à ontogénèse] Formation et développement des espèces vivantes au cours des temps; p. méton., étude de ce processus. À quelles conditions la phylogénie et l'ontogénie revêtiront-elles un caractère proprement scientifique (...)? (BLONDEL, Action, 1893, p.74). La phylogenèse a ses lois propres par rapport à l'ontogenèse, car dans le monde humain, l'ultérieur modifie la constitution de l'antérieur au moment de son apparition (J. VUILLEMIN, Être et trav., 1949, p.48).Prononc. et Orth.:[ ], [-
], [- -], [
-], [ ]. V. -
]. V. - . Étymol. et Hist. phylogénèse, phylogénie 1874 (E. HAECKEL, Hist. de la création des êtres organisés, trad. de l'all. par Ch. Letourneau, p.248, p.190 et 274). Comp. des élém. formants phylo- «espèce, classe organique», gr.
. Étymol. et Hist. phylogénèse, phylogénie 1874 (E. HAECKEL, Hist. de la création des êtres organisés, trad. de l'all. par Ch. Letourneau, p.248, p.190 et 274). Comp. des élém. formants phylo- «espèce, classe organique», gr. 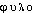 -, de
-, de 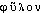 «classe, espèce» et -genèse, -génie, par l'intermédiaire de l'all. (E. HAECKEL, Generelle Morphologie der Organismen, II, XX, 299 et I, III, 57 ds NED Suppl.2). Fréq. abs. littér.:19.DÉR. Phylogénétique, phylogénique, adj. Relatif à la phylogénèse. Chez les Équidés et les Camélidés, on suit, de terrain en terrain, les étapes du progrès phylogénique (CUÉNOT, J. ROSTAND, Introd. génét., 1936, p.60). G. Roheim, qui substitue la théorie ontogénique de la culture à la théorie phylogénétique de la personnalité (Traité sociol., 1968, p.407). — [
«classe, espèce» et -genèse, -génie, par l'intermédiaire de l'all. (E. HAECKEL, Generelle Morphologie der Organismen, II, XX, 299 et I, III, 57 ds NED Suppl.2). Fréq. abs. littér.:19.DÉR. Phylogénétique, phylogénique, adj. Relatif à la phylogénèse. Chez les Équidés et les Camélidés, on suit, de terrain en terrain, les étapes du progrès phylogénique (CUÉNOT, J. ROSTAND, Introd. génét., 1936, p.60). G. Roheim, qui substitue la théorie ontogénique de la culture à la théorie phylogénétique de la personnalité (Traité sociol., 1968, p.407). — [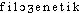 ], [
], [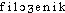 ]. — 1res attest. a) 1874 phylogénique (E. HAECKEL, op. cit., p.190), b) 1897 théories phylogénétiques (L'Année biol., 3, p.514); dér. avec suff. -ique de phylogénèse, phylogénie.phylogénèse [filɔʒenɛz] ou phylogenèse [filɔʒənɛz] n. f.ÉTYM. 1874, trad. de Haeckel; var. phylogénie, 1874, ibid.; d'après l'all., du grec phulon « race, tribu », et -génèse, -genèse.❖♦ Biologie.1 Mode de formation des espèces, développement, généalogie de l'espèce (par oppos. à ontogénèse; → Onto-, cit..)2 (XXe). Partie de la biologie qui traite de cette évolution. || La phylogénèse s'appuie sur l'étude des fossiles.❖DÉR. Phylogénétique.
]. — 1res attest. a) 1874 phylogénique (E. HAECKEL, op. cit., p.190), b) 1897 théories phylogénétiques (L'Année biol., 3, p.514); dér. avec suff. -ique de phylogénèse, phylogénie.phylogénèse [filɔʒenɛz] ou phylogenèse [filɔʒənɛz] n. f.ÉTYM. 1874, trad. de Haeckel; var. phylogénie, 1874, ibid.; d'après l'all., du grec phulon « race, tribu », et -génèse, -genèse.❖♦ Biologie.1 Mode de formation des espèces, développement, généalogie de l'espèce (par oppos. à ontogénèse; → Onto-, cit..)2 (XXe). Partie de la biologie qui traite de cette évolution. || La phylogénèse s'appuie sur l'étude des fossiles.❖DÉR. Phylogénétique.
Encyclopédie Universelle. 2012.